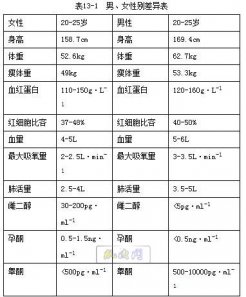
|
三周训练即可产生运动时胰高血糖素反应明显降低。在相同绝对负荷或相同相对负荷的运动时,训练后血中胰高血糖浓度比训练前为低。 2.胰岛素在进行递增负荷的运动中,随着动强度的增大和运动时间的延长,血中胰岛素浓度逐渐减低(图10-15)。血浆胰岛素的浓度在运动结束时,可下降到安静水平的50%以下。在2或3 h力竭性跑可能有更大的下降。

运动时血浆胰岛素下降的两个可能原因是: (1)运动时交感神经系统活动增强,儿茶酚胺对胰岛 B细胞的α受体的刺激比对 β受体的刺激更强烈, B细胞的分泌被抑制。 (2)运动时流经活动肌肉的血液明显增加,活动骨骼肌摄取胰岛素增加, (三)皮质醇 从轻度到中等强度运动时血液中皮质醇浓度的变化,文献报告不一。在进行剧烈运动时,血浆皮质醇浓度升高是肯定的(图10—16),这种升高可持续到运动结束后2 h。 David的研究表明,60%V02max的运动强度是引起血皮质醇浓度升高的强度阈。

低于60%VO2max的运动,使血浆皮质醇浓度下降。 剧烈运动时,血浆皮质醇浓度升高是由于分泌量的明显增加超过了它的清除量。这种分泌量的增加是由于剧烈运动时,体温升高和精神紧张等应激因素,对下丘脑的刺激增大的结果。 进行大强度长时间的力竭性运动时,血浆皮质醇的浓度反而下降(表10—2),这种现象是机体对极量运动的一种保护性反应。
有人观察了训练对安静时血浆糖皮质激素水平的影响,Frenk等发现,训练3周后安静时血浆糖皮质激素水平升高,训练6周后又回到训练前的水平。这反映了机体对来自内外环境的各种应激,逐步产生适应性反应, 训练后,可以进行相对强度相同的训练。 运动时的血浆促肾上腺皮质激素浓度升高的幅度比训练前大(图10—16)这种训练的效应反映了受试者训练后,要达到相同的相对负荷,必须以更大的绝对强度进行运动。
(四)生长素 生长素的释放量与运动的强度和持续时间有关。无训练男子在自行车功量计上,进行递增负荷运动时的研究发现,进行轻微运动和最大强度运动时血浆生长素水平均不升高,而进行中等强度运动时血浆生长素水平升高,从安静时的8.1 μg·L-1增加到13.6μg·L-1(图10—17)。

业已证明,运动强度必须达到临界水平才能引起生长素的分泌。 Metivier报告,高度训练的运动员在自行车功量计上进行50%VO2max的运动时,血浆生长素水平没有明显变化;当运动强度增加到66%VO2max时,血浆生长素水平明显升高。所以50—66%VO2max之间的某一点是引起生长素分泌的临界点。生长素最大反应的幅度和最大反应速率,取决于运动的强度。 BacKler发现,随着运动强度加大,运动25—60min后,血浆生长素含量达到高峰。如果继续加大强度,血浆生长素含量转向下落。 如果进行足以引起生长素分泌的中等强度运动,并持续一个当长的时间,结果出现生长素水平的稳定时相,这个时相一直维持到运动结束。 Hartley发现,一组缺乏训练的人以75%VO2max的强度在自行车功量计上进行40min运动,血浆生长素水平升高,若以此强度运动到力竭时,一部分受试者的血浆生长素水平下降到运动前水平。 在持续运动中,血浆生长素水平的升高有一个潜伏期。曾对一名35岁男子进行了多次实验,负荷强度由750kg m·min-1逐渐提高到1800kg m·min-1,而运动持续时间从180min逐渐缩短到2.5min,血浆生长素增多的潜伏期随之缩短,但均不短于10min。有研究报告提出, 有训练者运动时血中生长素含量的升高比无训练者低,训练后保持较高水平血浆生长素的时间较长。 Sutton等还发现,有训练者和无训练者在短时间运动时生长素都升高,但有训练者恢复到正常水平较快。 (五)甲状腺激素 一次急性运动后,游离甲状腺素可提高35%左右。游离甲状腺素浓度的升高是由于血浆蛋白与甲状腺素结合减少所致。训练导致安静时的甲状腺激素的总浓度稍下降,而游离甲状腺素的浓度稍升高。 Irvine用I131标记的甲状腺素注入血浆,证明运动训练能提高 T4的周转率,在训练期末运动员血浆 T4的半寿期为4.12天,而对照组的半寿期为6.93天。 二、运动时激素对机体代谢的调节 (一)激素对糖代谢的调节 1.激素对糖原分解的调节 ( l)儿茶酚胺。在运动过程中,肌糖原分解是依靠儿茶酚胺的作用。儿茶酚胺对糖原的分解作用主要是活化腺苷酸环化酶,增加 CAMP的产生,从而促进糖原的分解。 当用肾上腺素注入肌肉时, CAMP增加100%,磷酸化酶b的活性从占10%提高到40%,糖原合成酶I的活性下降,阻断狗的肾上腺素能神经的调节。血浆去甲肾上腺素减少,产生的乳酸也减少,无氧工作能力明显降低。切除肾上腺髓质的白鼠,运动中肌糖原的排空减慢,磷酸化酶( a十b)的活性降低,运动时儿茶酚胺分泌增加,将导致肌糖原分解,从而为运动提供更多的能量。肾上腺素对不同类型肌纤维的糖酵解的影响不同,对快肌纤维影响小,对慢肌的影响大。 例如,在跑前60min给动物皮下注射肾上腺素(0.1—0.5mg·Kg-1体重),跑后快肌中糖原没有完全耗尽,而慢肌和中间肌中糖原已所剩无几。可能是由于快肌纤维中毛细血管分布较慢肌少,从血液中获得肾上腺素较少。有实验证明,两种肌纤维对肾上腺素的敏感性是一致的。 运动时,儿茶酚胺也有促进肝糖原分解的作用,虽然肝糖原分解过程对儿茶酚胺的敏感性远不如胰高血糖素,但大强度运动时机体快速而大量释放儿茶酚胺也足以使肝糖原分解增加,使血糖明显升高。当鼠游泳60min而致低血糖后,注射肾上腺素可见血糖明显升高。Dnaish观察到鼠负重游泳100min时,切除肾上腺髓质的鼠,肝糖原减少的量少于正常鼠。另一个实验报告,当小鼠负重(体重的2%)进行力竭运动,在开始75min时,正常鼠的解糖原明显下降;切除肾上腺髓质鼠的肝糖原无明显变化,至力竭时,切除肾上腺髓质鼠比正常鼠的肝糖原高。二组动物达力竭的运动时间分别为191土8min和236士9min。 (2)胰高血糖素。胰高血糖素是肝糖原分解的强有力的调节因素。与儿茶酚胺一样,胰高血糖素的作用也是通过肝细胞膜上的胰高血糖素受体来促进肝糖原分解的。长时间的剧烈运动的,血浆胰高血糖素的升高,可以动员肝糖元的分解,在胰岛素浓度低下时,即使运动时胰高血糖素浓度不变,好对胰高血糖素会变得更敏感。 (3)性激素。用雌二醇处理阉割的雄鼠,在游泳的第 l h时肝糖原的动用不明显;在游泳到第5 h末时,低血糖发展仍不明显。说明雌二醇抑制糖元的分解。 2.激素对血糖利用的调节 (l)胰岛素。胰岛素是利用血糖的基本调节因素。血葡萄糖通过细胞膜进入肌细胞要由胰岛素进行调节。切除胰腺的狗运动时,血浆内葡萄糖不减低,注射胰岛素后,运动引起低血糖症。运动时,一方面是循环血中胰岛素浓度下降,但工作肌的毛细血管开放增加,血流量增加,进入工作肌的胰岛素总量增加,所以,有利于肌肉对血糖的利用;另方面,血浆胰岛素浓度下降,减少了非工作肌和其他组织对血糖的利用,有利于维持血糖的水平,保证神经细胞的糖供应。 (2)儿茶酚胺。肾上腺素促进肝糖元分解,使血糖升高。皮质醇和胰高血糖素促进脂肪的分解和利用,促进肝糖原分解作用。所以,运动时葡萄糖的利用增强,是通过肾上腺素、皮质醇和胰高血糖素的综合作用实现的。 3、激素对糖异生的调节 运动时,肝糖原分解加强的同时,常伴有肝糖原异生作用的加强,肝葡萄糖输出中的大约70—75%来自肝糖原的分解,其余由乳酸、丙酮酸、甘油和生糖氨基酸异生而来。长时间中等强度运动时,肝葡萄糖输出提高2倍,异生成的糖在肝葡萄糖输出中的比例由25%提高到45%,糖异生速率提高约3倍。加强糖异生作用的激素有糖皮质激素、胰岛素、胰高血糖素和肾上腺素。 ( l)糖皮质激素。在剧烈运动时,糖皮质激素能促进肝脏摄取更多的乳酸和丙酮酸等物质,加强糖的异生作用。在对鼠进行长时间游泳的研究中发现,运动开始后的策4 h肝糖原明显下降;当继续运动时,由于肾上腺皮质激素分泌的显著增加,并促进脂肪酸氧化和糖异生作用,所以在继续游泳到第5一12 h中肝糖原无进一步的变化;当游泳至第12h以后,血中肾上腺皮质激素低于运动井始时的水平,糖异生减弱,结果引起肝糖原第二次下降。 (2)胰高血糖素。运动时,皮质醇的升高促使丙氨酸和其它氨基酸从肌肉中释放出来,血液中丙氨酸浓度升高,从而加速葡萄糖一丙氨酸循环。长时间运动时,血浆胰岛素和血糖浓度降低,血浆胰高血糖素浓度升高,促使肝细胞摄取氨基酸异生肝糖元, (3)儿茶酚胺。剧烈运动时,肾上腺素浓度升高,能促进乳酸的糖异生。切除肾上腺髓质的鼠运动时,乳酸转变为葡萄糖的过程受抑制。 (4)胰岛素。运动时胰岛素分泌减少,葡萄糖磷酸激酶的活性下降,糖异生酶的活性升高,并且糖异生的底物如乳酸、丙酮酸等的含量也增加,从而糖异生加强。 (二)激素对脂肪代谢的调节 脂肪作为能源物质在人体内的贮量几乎是不受限制的,关键在于动用的调节。脂肪脂解产生的自由脂肪酸可被各组织利用,酮体(主要是 β—羟丁酸)可被中枢神经系统利用。 参与脂肪动用调节的激素,主要有儿茶酚胺、甲状腺素、腺垂体激素、糖皮质激素和胰岛素。 1.儿茶酚胺 运动时交感—肾上腺系统活动的加强,促进脂肪的脂解作用。肾上腺素能刺激激素敏感的脂解酶的活性,使动脉血中自由脂肪酸的浓度升高,使组织动用自由脂肪酸氧化供能,从而起到节省组织对血糖的利用,对维持血糖的相对稳定起着重要的作用。切除肾上腺髓质鼠运动时自由脂肪酸浓度升高推迟,正常鼠的脂肪酸则迅速上升; β—肾上腺素能阻断剂使狗长跑能力下降,这也与血浆游离脂肪酸浓度下有关。 β—肾上腺素能阻断剂也能降低骨骼肌中甘油三酯酶的活性。化学性阻断鼠的交感神经节后纤维,当鼠跑至力竭时,血浆游离脂肪酸水平比正常鼠低得多,跑至力竭的时间也短(正常鼠为121土135,阻断鼠为42士6S) 2.甲状腺激素 狗经甲状腺素处理后,进行15、30和60min跑步时,游离脂肪酸的动员增多。甲亢者运动时,血浆游离脂肪酸浓度的升高比正常人多。切除甲状腺的狗运动时,血浆游离脂肪酸浓度和周转率远远低于正常狗。 3.肾上腺皮质激素 切除肾上腺皮质的动物长时间运动时,血浆游离脂肪酸水平降低,但先给予强的松后,这种变化可以逆转。 性激素在一定程度上也参与脂解反应的调节,切除卵巢并用雌激素处理的雌鼠在跑台上跑至力竭时,血浆甘油三酸的降低比正常雌鼠明显。这是由于雌激素加强脂蛋白脂酶活性,提高心肌等对脂肪酸的利用能力。 4.腺垂体激素 生长素促进肌中甘油三脂的氧化,并使肌组织的甘油三酯贮量减少。运动时,血浆生长素浓度升高常伴随游离脂肪酸升高,切除垂体的动物运动时,游离脂肪酸水平的升高受到抑制。但从人体得到一些结果与上述动物实验中的结果是矛盾的,所以腺垂体激素对人体运动时的脂解作用的调节尚待进一步研究澄清。 5.胰岛素 胰岛素有抗脂解的作用。在长时间运动时,血浆胰岛素水平的降低是由于提高脂肪脂解的过程,进而促进游离脂肪酸的氧化。 切除胰腺的狗在25min剧烈跑时,血浆游离脂防酸浓度升高,而对照狗在同样25min跑时,血浆中游离脂肪酸浓度减低。长时间运动时,血浆胰岛素浓度降低导致游离脂肪酸氧化加剧,使产生的乙酰辅酶 A增多,此时乙酰辅酶 A将聚合成酮体(如 β羟基丁酸、丙酮等),但肌肉和脑组织可以利用酮体物质作为燃料,放一般说来运动时血酮浓度不会有急剧的升高。 (三)激素对运动后恢复期糖代谢的调节 1,儿茶酚胺 交感—肾上腺系统在动用糖贮备的过程中发挥作用。运动结束后,血中儿茶酚胺浓度迅速降低即有利于糖贮备的恢复。运动后即刻注射肾上腺素,糖原恢复和乳酸消除的速度减慢。 2.胰岛素 在糖原合成中,糖原合成酶是一个关键酶。肌肉和肝内的儿茶酚胺和胰高血糖素可激活糖原合成酶激酶,此酶催化有活性的糖原合成酶 I转变为无活性的糖原合成酶 D。胰岛素的作用相反,降低糖原合成酶激酶的活性,使糖原合成酶D转变为糖原合成酶 I。 3.皮质醇 皮质醇能诱导肝内6-磷酸葡萄糖酶和其他糖异生酶的生成。因此在运动结束后的恢复期,皮质醇对糖原的恢复起一定的作用。给切除肾上腺的鼠注射糖皮质激素,运动结束后恢复期内肝糖原的恢复比对照组快。运动后血浆皮质类固醇升高伴有心肌糖原的超量恢复。当心肌糖原超量恢复时,骨骼肌糖原也恢复到运动前的水平,继而骨骼肌中的糖原贮量也出现超量恢复。 4.性激素 阉割鼠游泳 5h后,给予雌二醇时,心肌糖原的超量恢复的幅度增大。给阉割动物注射睾酮,也可增加肌糖原合成。这些说明,性激素有促进糖原贮备恢复的作用。 |


“肌肉网”

“健身餐食谱”

“教练库”