|
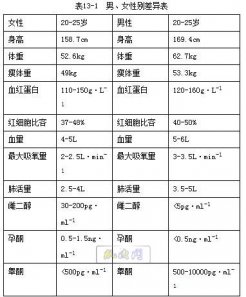
表明膜内、外两侧存在90mV的电位差,且细胞膜外是正电位。如规定膜外电位为零,即膜内电位为—90mv。 由于这一电位差是存在于细胞膜两侧的,故称跨膜静息电位,简称为静息电位或膜电位。 只要细胞未受到外界刺激,静息电位就稳定在某一固定水平。细胞安静时,膜内、外电位稳定于某一数值的这种状态,也称做极化状态。 静息电位的成因是由于细胞膜内外 Na、 K离子分布不均和细胞膜具有选择通透性。 在静息状态下,由于膜内的 K+浓度比膜外的高20-40倍,膜外的Na离子浓度比膜内的高7-12倍 故在膜两侧存在着Na、 K离子的浓度梯度。这一驱使K离子顺该梯度向膜外扩散, Na顺浓度梯度向膜内扩散;另一方面,由于细胞膜对 K+的通透性要比对 Na的通透性大20一100倍左右,故安静时,细胞膜对 K+有一定的通透性(虽然很低),而对Na基本是不通透,从而使少量的 K+通过细胞膜扩散到膜外,而Na+则不能扩散到膜内。同时,由于细胞内负离子大多是大分子的有机磷酸离子和带负电荷的蛋白质,不能随 K一道通过细胞膜,而留在膜内。这样一来,细胞膜外侧就有较多正离子,细胞膜内侧有较多负离子。因而使膜外电位变成正,膜内电位变成负,使膜内外产生了外正内负的跨膜电位差。 可见这种跨膜电位差是由于膜内的K+向膜外扩散而形成的。 一当由 K外流形成能这种内负外正的电位差即电位梯度出现后,它就成为一种阻止 K+继续向外扩散的力量。因之,当由于浓度梯度而使 K+向外扩散的力量和电位梯度阻止 K+向外扩散的力量相等时,即两种力量达到平衡时, K+就不再继续向外扩散;膜两测的电位蓑也就稳定于某一固定数值。这就是静息电位。 各种动物细胞都有外正内负的静息电位,但由于各类细胞膜内外 K浓度比值或膜对 K的通透性不同,因而其静息电位窿也有所不同,例如,哺乳动物的心肌,骨路肌和神经细胞的静息电位为-70一-90mV,平滑肌为-5O一-60mV。 (二)动作电位 在神经纤维的左测给予一个短促而有足够强度舱刺激时,它就产生一次兴奋,当兴奋传至微电极处时,就可在示彼器上观察到一次电位波动,即动作电位。此时,膜内原存在的负电位迅速消失并进而变为正电位,由原来的—90mV迅速升高为十30mV左右,即出现膜电位的倒转。由静息电位时的外正内负转变为内正外负,膜内电位由—90mV上升到0mV,为负电位的消失。膜内负电位的消失称为去极化。再由0mV上升十30mv左右称为超射。故整个膜内、膜外电位变化的总范围是120mV左右,这就是的动作电位幅度,并构成动作电位的上升支,也叫除极化时相。动作电位在神经纤维上只持续0.5-1ms,当上升支迅速达到高峰后,立即迅速下降到原来的静息电位水平 这就构成动作电位的下降支,也叫复极化时相。动作电:位曲线很象一个尖锋,故又称锋电位。在用高倍放大和慢扫描所得的记录上,在锋电位之后还常常可看到一个振幅低但持续时间很长的后电位(图 l—4)。 锋电位在刺激后几乎立即出现,潜伏期不超过0.06ms,锋电位持续时间约0.5 ms。其中绝对不应期相当于从触发动作电位起到完成复极1/3的时间,从这一点向后到负后电位的开始,即相对不应期(图 l—4)。每一刺激引起一个锋电位,锋电位的传播速度与神经冲动传导速度相等,故锋电位就是兴奋过程或冲动。继锋电位而出现的后电位则与兴奋后的恢复过程有关。 二、静息和动作电位的成因 (一)静息电位成因如前所述 (二)动作电位的成因 概括地说,是在刺激的作用下,细胞膜保持的极化状态逐步被毁除,称为除极; 当除极化达到临界水平时,即膜电位下降到-50一-70 mV水平时,就触发了动作电位,这个能触发动作电位的膜电位就叫阈电位。 引起阈电位的最小刺激是阈刺激,其强度值就是阈值。当膜电位降低到阔电位子当膜电位降低到阈电位水平时,细胞膜的通透性突然改变,膜对 Na+的通透性突然增大约500,大大超过对 K+的通透性。 Na+由于浓度梯度(膜外助 Na+浓度比膜内高12倍)的推动和膜内负离子的吸引而迅速内流,使膜内正电荷迅速增加,从而引起两侧电位由安静时的内付外正倒转为内正外付, 也就形成了动作电位的上升支既除极化时相。但是膜对Na的通透性增加只是暂时的,当除极化达到顶点时,由Na离子造成的膜内正电位阻止Na离子的内流,膜对的通透性降低,而对K离子的通透性升高,这时膜又几乎只对K有通透性,于是 膜内的K离子由于高浓度和正电位的推动而迅速外流,使膜内外电位又恢复到内付外正状态, 形成动作电位的下降支,既复极话时相。由此可见,动作电位曲线的上升支,是N内流所形成的,但当 N内流所形成的膜内正电位所产生的电位梯度与推动 Na+内流的浓度梯度相平衡时,Na+就停止继续流入,除极化达到顶点,这时膜两侧的电位差就是动作电位的高度。 根据动作电位成因的分析,可以说明各类可兴奋细胞动作电位的某些共同特性,例如,不论使用何种性质的刺激,只要达到一定强度,它们在同一细胞所引起的动作电位的波形和变化过程都是一样的,并且在刺激强度超过阈刺激以后,即使再增加刺激强度,也不能使动作电位的高度进一步加大, 这个现象,称做“全或无”现象。这是因为,动作电位只是由阈电位所触发,至于动作电位所能达到的大小,主要决定于当时膜内外 Na+、K+的浓度比而不决定于刺激的强度。 三、动作电位的传播 动作电位的特征之一就是它的可传导性,即细胞膜任何一处所产生的动作电位,可传播到整个细胞。其机制是:当膜某一点受到刺激而 产生动作电位时,该点的膜电位即例转为外负内正,而邻接的末兴奋部位因仍处于静息状态,膜电位为外正内负,再由于细胞内液和细胞外液都是导电的,于是兴奋部位与末兴奋部位之间就产生局部电流。 这一局部电流构成了对未兴奋部位的刺激,使之产生兴奋,就象外加刺激在最初的受刺激部分引起兴奋一样。这个过程在膜上连续进行下去,表现为动作电位在不断向前传播,直至传遍整个细跑。在传播中,它的前部不断地向前传导,它的后部又不断复极化,使局部电统波及的长度不变。 由于兴奋本身就是动作电位,因之,兴奋在同一细跑上的传导,也就是动作电位的传播。在神经纤维上传导着的兴奋或动作电位,称做神经冲动或简称冲动。正如超过阈值的强度不影响它所引起的动作电位的大小一样,动作电位在单一细胞上的传导距离和传导速度也不决定于最初刺激的强度,动作电位的大小也不会因传导距离的增大而有所减弱。此外,如刺激作用于细胞约中段,由此而产生的动作电位可同时向两端传导,这是由于局部电流可以向两侧传导的缘故。在完整的反射弧中,兴奋只能沿单一方向传导,这是由于反射弧各环节衔接处,兴奋只能沿单一方向传递而不能逆传。因兴奋在反射弧各衔接处的传递,起作用的除锋电位外,还有更复杂的化学过程参与。

四、局部兴奋 刺激强度小于阈刺激,虽然不引起可传播的动作电位,但并非对细胞不产生任何影响。 实验表明,阈下刺激可使受刺激局部对 Na+的通透性轻度增加,造成原有的静息电位的轻度减小,并且只局限在受刺激的局部范围,这种电位变化,称为局部兴奋。 局部兴奋的生理意义在于它可以总和。由于局部兴奋不是“全或无”式的,而是随刺激强度的增加而增强,且没有不应期,故在第一个阈下刺激所引起的局部兴奋尚未消失以前,紧接着再给予另一个刺激,两个刺激所引起的局部兴奋就有可能叠加起来使膜电位降低到阈电位水平,而触发可以传导的动作电位,称为时间总和。同样,相邻的细胞膜同时受到两个阈下刺激时,这两个刺激所引起的局部兴奋也可以总和起来,称空间总和。时间总和多见于感受器、神经肌肉接头等处,空间总和多见于中枢突触部位。 |


“肌肉网”

“健身餐食谱”

“教练库”