|
第一节吸氧量和氧亏 一、需氧量、吸氧量与耗氧量以及氧亏与氧债 (一)需氧量是指人体单位时间内所需的氧量。 成年人静息时为了维持各组织器官的基本生理活动,约需氧250ml/ min或3.5mI氧·/kg 凡运动强度愈大,需氧量就愈大,如100 m赛跑的速度计算成每分需氧量可达4OL/min,而中等强度的马拉松跑时的需氧量约为2—3.5L/ min。运动时的绝对(总)需氧量则随运动持续时间而异。如100 m赛跑历时仅12s左右,其总需氧量竞达7 L左右;持续时间达2 h以上的马拉松跑,其全程的总需氧量达700 L以上。人体单位时间内(通常以 lmin计)从肺泡气中获取的氧量或全身各组织器官从毛细血管中抽提的氧量称为吸氧量或每分吸氧量(VO2)。人体单位时间所消耗的氧量称耗氧量 (二)氧亏与氧债 人在从事持续时间短、强度大的运动项目,即高功率项目的运动时所需要的氧量极大。 例如,用12 s(或更短的时间)跑完100 m,根据计算,需要7 L氧,折合为35L/min、(580ml/s),正常人为250ml/min(4.17ml/s)假如人体能从动脉血中摄取180ml/L氧,那么心脏需要输出200L/min血量才能满足需要,然而人体心脏无论如何不可能达到如此巨大输出量的。因此,在这种性质的运动时即使氧运输系统的功能能力动员到极限,其供氧量仍远远不能达到需氧量的水平。这时需氧量与供应量之差称为氧亏。在适度的运动中,需氧量虽然在人体氧运输系统购贮备力的范围内。例如,一般大学生以75s跑400 m的速度能跑完3000 m,其需氧量约为1.0一1.5L/min。但由于人体的氧运输系统功能的惰性,在运动的开始阶段,氧运输系统的功能并不能立即提高到与运动的需要相适应,所以在运动的开始阶段也有氧亏出现

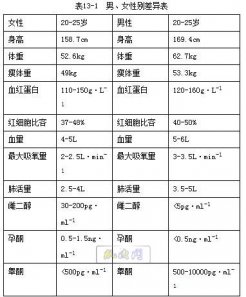
运动后恢复期内的过量氧耗就是用于偿回运动过程中的氧亏,所以把运动后恢复期内的过量氧耗称为氧债。 经典的氧债学说认为, 1.快恢复时相是偿回非乳酸成分的氧债,即这部分过量的氧消耗是用于重建和恢复运动时消耗的 ATP和 CP贮备的,而 ATP和 CP的消耗并不伴有乳酸的产生和积累,故称为非乳酸氧债。 2.慢恢复相的过量氧耗,是用于清除运动过程中糖原无氧酵解生成的乳酸,故称为乳酸氧债 人体负氧债的能力随训练水平的提高而增强,一般人在剧烈运动中约能负10L—氧债,其中25%属非乳酸氧债、75%属乳酸氧债。 受过良好训练的人,负氧债能力的上限可达15—20L 二、运动后的过量氧耗 (一)运动后过量氧耗的生理基础 近年来的研究对经典氧债学说提出了挑战, Brooks等指出,运动后出现的过量氧耗,并不是完全用于偿回运动中的氧亏,而是由于运动时激活了肌细胞中线粒体氧化代谢,使之处于高代谢水平上;在运动结束之后的一段时间内,许多激活线粒体酶的因素并未立即消除。例如,肌肉的温度、血液中儿茶酚胺 甲状腺素和去甲肾上腺索的浓度等因素,在运动结束后均未立即恢复到静止水平。实验证明,给狗注射去甲肾上腺素,其骨骼肌肉的耗氧量增加40%;现已证明,肌温是运动后耗氧量保持较高水平的主要原因之一。实验表明,肌温.与氧耗的恢复曲线是同步的。据估算,温度的代谢效应可能占运动后恢复期耗氧量慢成分的60一70%。根据近代的研究材料, Brooks建议用“运动后过量氧耗”(EPOC)的术语来代替“氧债”这一名词,因为氧债并不能反映运动后氧耗水平处于较高水平的真正机制。反而引起一种错觉,认为适动后恢复期内耗氧量之所以较高,纯粹是由于偿回运动过程中欠下的债务所致。 运动的净需氧量=[运动其总吸氧量+恢复其总吸氧量]-[恢复时间+运动时间]×250ml (二)运动后过量氧耗的影响因素 1.儿茶酚胺的影响 剧烈的运动使体内儿茶酚胺浓度增力口,而运动后恢复期儿茶酚胺的浓度仍然保持在较高水平,因而氧消耗增加。格莱登(Glader)的实验证明,给狗注射去甲肾上腺素,狗的骨骼肌的耗氧量增加40%。 2.甲状腺和糖皮质激素的影响 甲状腺和糖皮质激素也促进细胞膜上的钠钾(Na+—K+)泵的活动增强。同时,在运动后恢复期,其浓度仍然保持在较高水平,因而消耗一定的氧。 3.体温升高的影响 海克勃(Hegberg)的研究指出,在运动后恢复期的耗氧量成分中,恢复曲线的慢成分有60%—70%产生于肌肉温度升高的原因。实验证明,体温和肌肉温度与运动后恢复期耗氧量的恢复曲线是同步的。因此,在运动后恢复期体温不可能立即恢复到安静时的水平,使肌肉的代谢继续维持在一个较高水平上。 第二节有氧工作能力的评价 一、最大吸氧量及影响因素 (一)最大吸氧量(VO2max) 是指人体在进行有大量肌肉群参加的力竭性运动中,当氧运输系统中的心泵功能和肌肉的用氧能力达到本人的极限水平时,人体每单位时间(通常以 min为单位)所能摄取的氧量。 所以最大吸氧量(maximal oxygen consumption VO2max)是评定人体有氧工作能力(aerobic working capacity)的综合指标。 (二)最大吸氧量的表示方法与参考值 最大吸氧量的表示方法,有绝对值与相对值两种, A.前者是表示整个机体在单位时间(lmin)内所能吸取的最大氧量,我国青年学生的最大吸氧量约为3.5L /min但由于个体的身高;体重等差别甚大。 B.运动生理学中常以人体的相对值来表示,最常用的是按每千克体重计的最大吸氧量(ml/ kg.min)。一般说来,我国男大学生的最大吸氧量在50—55ml/kg.min之间,女大学生约比男学生低10%左右,最大吸氧量在少儿期间随年龄的增长而增加。 青春发育期前,男女性别之间无差别,女子12岁时开始性发育,便开始出现性别差异。以绝对值计的最大吸氧量,其性别差异可达10%以上,若以每千克体重计,则性别差异减小。 女子最大吸氧量(ml/ kg/min)比男低的原因有下列几点: ( l)女子的心室容积小于男子;故泵血量较男子少; (2)女子的血‘红蛋白总量较男子低,放血氧容量低于男子。 (3)女子的脂肪较男子多,故相对的肌肉重量较男子为少。 男子在18—20岁时最大吸氧量值达峰值,女子在14一16岁时即达峰值。男子最大吸氧量的峰值能稳定的保持到30岁左右,女子约稳定的保持到25岁左右。以后,最大吸氧量将随年龄的增加而递减,这种自然减退的进程,估计大约每10年降低9%左右,即70岁老人的最大吸氧量相当于其本人青年时值的64%。如若坚持体育锻炼,最大吸氧量随年龄而降低的幅度可以减小。 纵向与横向的研究材料都表明,耐力训练可促进最大吸氧量的增长。据国外报道,男子越野滑雪运动员最高的最大吸氧量值达94ml/kg.min,女子运动员最高值达85.1 ml/ kg.min,Bergh等(1978)的横向研究也表明,耐力性项目如越野滑雪运动员和长跑运动员的最大吸氧量最大,短跑运动员的最大吸氧量最小
 
二、影响最大吸氧量的因素 (一)决定最大吸氧量的生理机制 A.训练对最大吸氧量中央机制的作用 心泵功能强弱的指标是运动时心脏的最大心输出量的大小,而最大心输出量则是最高心率与最大搏出量的乘积。为了阐明训练影响对最大吸氧量中央机制的作用,必须分别探讨训练对HRmax和 SVmax的影响。 1.据报道,优秀的耐力运动员的静息心率仅为40次/ mig左右;但训练对个体的 HRmax没有影响。有一部分学者还认为,耐力训练可导致 HRmax减少3—5次/mis。可见,训练致使极量运动中 COmax增加,并不是由于HRmax增加。 2.SVmax的大小是最大吸氧量中央机制的又一个重要方面。SVmax的大小决定于心脏容积和心肌收缩力。有许多研究表明,耐力训练可使心室腔增大,、常人的心容积为700—800ml(10-llml·/kg体重) ,而耐力运动员可达900一1000ml(14ml/Kg),耐力训练引致心腔增大为增进 SVmax提供了形态学基础。据文献报道,国外个别卓越的耐力运动员的 SVmax可达16Oml以上,其VO2max可达5L/min。 B.训练对最大吸氧量外周机制的作用 1.肌纤维的类型 对肌纤维类型的研究表明,慢肌纤维具有丰富的毛细血管分布,具有数量较多和体积较大的线粒体,线粒体中氧化酶系的活性也高,慢肌纤维中肌红蛋白含量也较高。以上这些特征都有利于增加肌纤维的摄氧能力,慢肌纤维的百分组成与最大吸氧量有密切的关系 2.肌肉的供血量 肌肉的供血量除了与心泵功能(心输出量属中央机制)有关外,还与血液的重新分配有关。 Rowell(1974)指出,在最大运动时,即使不考虑心输出量的增加,仅仅从腹腔内脏和肾血管的收缩,每分钟就可腾出2.2 L血液分配到活动肌肉中去,这样可使吸氧量增加500ml/min左右。训练导致心血管家统的调节功能的改进,有利于运动时肌肉获得更多的血液供应,从而使最大吸氧量得以提高 3.循环血量和血红蛋白总量与氧的运载量密切有关,所以与最大吸氧量的值也密切相关。前已提到,女子最太吸氧量的值所以比男子低,其中循环血量与血红蛋白总量较男子少是其原因之一,训练能否改变(增加或减少)循环血量与血红蛋白总量尚未见报告,预计训练对这两个指标不会有明显的变化,所以不能把它看作是训练引起最大吸氧量增进的机制。
 
(二)遗传、年龄、性别和训练对最大吸氧量的影响 (1)遗传因素 VO2max受遗传因素的影响较大。许多学者的研究也指出,VO2max与遗传的关系十分密切,其可训练性即训练使VO2max提高的可能性较小,一般为20%-25%。 (2)年龄、性别因素 VO2max在少儿时期随年龄增长而增长,并于青春发育期出现性别差异,男子一般在18-20岁时最大?氧量达峰值,并能保持到30岁左右;女子在14-16岁时即达峰值,一般可保持到25岁左右。以后,VO2max将随年龄的增加而递减。 (3)训练因素 长期系统进行耐力训练可以提高VO2max水平,戴维斯(Davis)对系统训练的人进行了研究,受试者的VO2max可提高25%,表明经训练VO2max是可以得到一定程度提高的。越野滑雪和长跑等耐力性项目的运动员最大?氧量最大,明显高于在非耐力性项目运动员和无训练者。 在训练引起VO2max增加过程中,训练初期VO2max的增加主要依赖于心输出量的增大;训练后期VO2max的增加则主要依赖于肌组织利用氧的能力的增大。但由于受遗传因素限制,VO2max提高幅度受到一定制约。 二、有氧耐力 有氧耐力(aerobiccapacity)是指人体长时间进行有氧工作的能力(糖、脂肪等有氧氧化供能)。 在超过30分钟的长时间的运动中,最大摄氧量不易出现,运动被维持在次最大摄氧量水平。例如,最大摄氧量平均为71.5 ml/kg·min的5名男子中长跑运动员,在1 500米、5 000米、10000米、20千米、30千米及42千米不同距离的跑动中,随距离的延长运动中的最高吸氧量下降。在实验室条件下进行5千米的跑台运动,优秀运动员的最高吸氧量水平可维持83.2%y(),。+他们的平均值在82.5—83.7 mi/kg·rain的范围内。在10千米的测试中,瑞典的滑雪运动员为87.5(79-92)%Vo2max,日本的运动员可达80%.最大摄氧量即使相同,运动成绩并非相同。也就是说,维持最高摄氧水平的能力不同。前世界马拉松竞走冠军的Vo2max为69.7 mi/kg·min,而他在跑中的强度是86%Vo2max。一般马拉松运动员的Vo2max。为70—80mi/kg·min范围,在跑中却是70%—80%,维持本人最大摄氧量的百分率,也是有氧耐力的一个重要因素。以最大摄氧量的90%、80%、70%的速度进行长跑运动时,也会出现个体差异。由此表明,有氧运动能力不仅与最大摄氧量的大小有关,而且与维持最高摄氧水平的能力有关。 (一)有氧耐力的生理学基础 空气中的氧通过呼吸器官活动吸进肺,与肺循环毛细血管之间进行气体红细胞中的血红蛋白携带,并运输到肌肉组织附近;红细胞释放出氧并弥散进入肌肉组织;肌肉组织利用氧进行有氧代谢活动。因此,影响有氧耐力的水平,主要取决于心肺功能、骨骼肌的特点、神经调节能力以及能量供应特点。 目前研究认为,心肺功能是影响VO2max的中枢机制,而肌纤维类型的百分比组 成及其骨骼肌的代谢特征是影响VO2max的外周机制。 1.心肺功能 肺的通气与换气机能影响人体吸氧能力。空气中的氧通过呼吸器官活动吸进肺,与肺循环毛细血管之间进行气体交换,肺通气量越大吸人体内的氧就越多,呼吸频率和呼吸深度影响肺通气量的变化。因此,运动时提高和掌握有效的呼吸动作,增强呼吸机能就可以提高有氧耐力。 心脏的泵血功能与有氧耐力密切相关。心输出量受每搏输出量和心率的制约。运动训练使心脏的形态与机能出现适应性变化。主要表现为左心室内腔扩张,心容积增大,安静时心率减慢,每搏输出量增加。而每搏输出量决定于心肌收缩力量和心室腔容积的大小。 红细胞的数量是影响有氧耐力的一个因素。血液中红细胞所含的血红蛋白携带氧进行运输。运动员血红蛋白含量假如下降10%,则往往会引起运动成绩下降。 2.骨骼肌的特征 肌组织的有氧代谢机能影响有氧耐力。肌肉内毛细血管网开放数量的增加,可使单位时间内肌肉血流量增加,血液可携带更多的氧供给肌肉。每100毫升动脉血流经组织时组织利用氧的百分率称为氧的利用率,表示肌肉利用氧的能力。肌组织利用氧的能力主要与肌纤维类型及其代谢特点有关。优秀的耐力运动员慢肌纤维百分比高,肌红蛋白、线粒体和氧化酶活性高、毛细血管数量多。 3.神经调节能力 大脑皮质神经过程的稳定性,以及中枢之间的协调性影响有氧耐力。长期耐力训练可以改善神经的调节能力,节省能量消耗,保持较长时间的肌肉活动。 4.能量供应特点 糖和脂肪在有氧的条件下,能保持长时间供能的能力是影响有氧耐力的重要因素之一。耐力性项目运动持续时间长,强度较小,主要以有氧供能为主。在运动中随着运动时间的延长,脂肪供能比例逐渐增大,糖原的利用减少。供能物质的储存、肌肉有氧氧化过程的效率、各种氧化酶的活性,以及动用脂肪 供能的能力,都可以通过有氧耐力训练来提高。 (二)影响有氧耐力的因素 有氧耐力的生理学基础均为影响有氧耐力的因素。最大摄氧量是有氧耐力的基础,其值越大,有氧耐力水平越高。肌纤维类型的百分组成、肌糖原的衰竭、运动中大量水分的丢失、肌细胞膜电解质平衡紊乱以及有氧氧化酶的活性等因素与有氧耐力水平有关。 三、乳酸阈与通气阈 (一)乳酸阈与个体乳酸阈 在有氧供能的渐增负荷运动中,运动强度较小时,血乳酸浓度与安静时的值接近,但是,随着运动强度的增加,乳酸浓度会逐渐增加,当运动强度超过某一负荷时酸浓度急剧上升的开始点,称为乳酸阈。乳酸阈是反映人体的代谢供能方式由有氧代谢为主开始向无氧代谢为主过渡的临界点,通常情况下,血液乳酸浓度为4 mmol/L,大约为最大摄氧量的60%~80%。通常用2 mmol/L或4 mmol/L(或称为血乳酸急剧堆积的开始点:OBLA)。由于个体的差异比较大,乳酸阈值并不都是4 mmol/L,其变化的范围大约在1.4—7.5 mmol/L之间,因此,也有称乳酸阈为“个体乳酸阈”。

最大摄氧量反映人体在运动时所摄取的最大氧量,而乳酸阈则反映人体在渐增负荷运动中,血乳酸浓度没有急剧堆积时的最大摄氧量实际所利用的百分比,即最大摄氧量利用率(,呖食02一)。其值愈高,有氧工作能力愈强;反之,有氧工作能力愈低(图13—4)。乳酸阈是反映有氧耐力的千个指标,也是有氧代谢能.力的组成部分。 (二)乳酸阈的生理机制及其影响因素 1.乳酸阈的生理机制 (1)运动时肌肉缺氧:人体在运动中随着运动强度的增大,运动肌肉的氧供应不足,使得一部分肌糖原在无氧的条件下,分解供能产生乳酸。肌乳酸由肌细胞扩散进入血液中,导致血乳酸浓度增高。 (2)需氧量大于机体的摄氧量:以有氧供能为主转向以乳酸供能为主,有氧供能逐渐减少,乳酸供能增加。 (3)肌纤维类型的动用:在低强度的运动中,慢肌纤维的动用占优势。随运动强度的渐增,快肌纤维的动用逐渐转向优势i导致血乳酸浓度增加。 (4)肝脏对乳酸的消除能力降低:在运动中由于血液的重新分配,流人肝脏的血液减少,降低肝脏郴L酸的消除能力。 (5)血乳酸浓度也与能量代谢物质的动用有关:在运动前大量摄取咖啡或高脂肪膳食,使血液中的游离脂肪酸浓度增加,在运动中脂肪作为能量物质的动用会抑制乳酸供能。 2.影响乳酸阈的因素 (1)性别、年龄的影响:性别影响乳酸阈时的吸氧量水平,但不影响乳酸阈时的最大摄氧量利用率百分比。研究表明,我国男大学生乳酸阈时的吸氧量比女子显著性增大,而吸氧量利用率百分比在男女之间却没有显著性差异。我国男女优秀运动员的最大摄氧量有显著性差异,乳酸阈时的吸氧量占最大摄氧量百分比却没有明显差异。年龄对儿童少年的乳酸阈有一定的影响。 斯金纳(Skinna)等研究认为,儿童时期由于果糖磷酸激酶水平低,所以用 4mmol/L的乳酸阈研究是没有意义的。许多学者对儿童少年进行研究后表明,9-15岁的乳酸阈在3.84—4mmol,/]5之间,与成人比较没有显著性差异,但5岁儿童乳酸阈值仅有2mmol/L。 (2)肌纤维类型及酶的活性:慢肌纤维百分组成高的人,其乳酸阈也高。泰斯克(Tesch报告了乳酸阈4mmol/L时的吸氧量与慢肌纤维之间的相关系数r:0.75。有氧耐力训练可提高氧化酶的活性。由此表明,乳酸阈的提高与肌纤维类型的动用、酶的活牲有密切相关,训练的目的就在于改善这些因素。 (3)训练水平的影响:最大摄氧量受遗传因素的影响,而训练可以提高乳 酸阈。戴维斯(Davis)的研究指出,经过系统训练后的受试者,最大摄氧量只能提高25%,而乳酸阈却可以提高44%。其原因是遗传因素限制了最大摄氧量的提高幅度,而乳酸阈值主要与外周的代谢因素的关系更密切,如肌肉的血流量、肌纤维类型的百分组成及酶的活性等。训练可以改善代谢能力,使乳酸阈值较大幅度的提高。 (4)运动项目的影响:乳酸阈值与耐力性运动成绩呈高相关,最大摄氧量固然重要,但与3 000米、5 000米、10000米及马拉松等长距离和超长距离的运动成绩都是中等的相关关系,而血乳酸与运动成绩具有重要的关系。由于在这种强度的运动中血乳酸浓度低,运动成绩主要依存于血乳酸浓度的减少和乳酸阈值的提高因此,长跑、游泳、自行车等项目运动员的乳酸阈值及吸氧量利用率百分比要高于短跑、短距离游泳等项目运动员。

(5)环境条件的影响:人在诲拔4000米的高处时,大气压的下降伴随着氧分压的减少,吸氧量也大为减少i同时也影响乳酸阈。高原条件下乳酸阈时的吸氧量明显低于平原地,温度的变化也影响乳酸阈。研究表明,高温(40摄氏度)条件下进行渐增负荷运动与常温(’25摄氏度)相比,乳酸阈时的吸氧量有明显的差异。 (三)通气阈及其产生机制 1.通气阈(ventilatory threshold,VT)

在递增负荷运动中,用通气变化的拐点来测定乳酸阈,称为通气阈。研究表明,在渐增负荷运动中,气体代谢的各种指标随运动强度的增加而逐渐发生有规律的变化,当血乳酸急剧增加时,通气量、二氧化碳排出量等指标发生非线性的上升。可用这种变化的特点来判断乳酸阈的发生。通气阈是判断乳酸阈的工种非损伤性的方法,其判断方法多采用通气量急剧上升的开始点来确定.但大多数研究者认为,采用气体代谢的各项指标进行综合性判断比较客观。 2.通气阈产生的机制 缺氧是引起通气量急剧增加的二个因素。运动强度缓慢地增加时,由于这种强度比较低.运动主要是有氧供能。随运动强度增大,有氧代谢产生的能量满足不了机体的需氧量,糖酵解代谢供能的比例增多,而使血乳酸浓度增加。体内碳酸氢盐缓冲系统,生成乳酸钠和碳酸,使细胞中的二氧化碳的产生量增加。这样,在有氧代谢所产生的二氧化碳量中又增加了一种由重碳酸钠缓冲而产生的二氧化碳量。由于动脉血中的HC03—减少,PC02和H+浓度增加。并刺激了颈动脉体化学感受器及呼吸中枢,为了维持体内正常的酸碱平 衡,排除更多的二氧化碳量而使通气量增强,产生了过多通气。因此,在乳酸阈时出现了通气量、二氧化碳排出量非线性增加、二氧化碳浓度下降现象。 (四)研究乳酸阈、通气阈的意义 乳酸阈、通气阈与运动训练、运动成绩有着密切的关系。这些指标也和最大摄氧量一样,被科研工作者利用于评价运动训练、运动成绩以及体力测定。同时在临床医学上利用通气阈及低强度的乳酸阈(1毫摩尔)来进行康复运动和为一般人制作运动处方提供依据。 1.评定有氧耐力 最大摄氧量代表人体的最大有氧能力。可是达到最大摄氧量时,人的心率为180次/分以上,呼吸商超过1.2以及血乳酸在94mmol/L以上。此时的运动能力可以说是人的运动能力达到了极限,即将停止运动。乳酸阈表示并未发生血乳酸急剧的增加而进行的最大运动能力(利用最大摄氧量的能力)。这种运动的主要能源来源于有氧代谢。 麦得(Mader)等研究者提出,用4mmol/L的乳酸阈时的跑速作为评定运动员有氧耐力的指标,相当于吸氧量的80%左右。乳酸阈值越高(强度或速度),利用有氧代谢的运动能力也就越大。由于乳酸阈反映肌肉的氧化能力,它与肌纤维中线粒体的数目、体积、氧化酶的活性以及毛细血管的发达密切相关。因此,可用乳酸阈评定运动能力和训练强度。 2.训练强度的制定 利用乳酸阈制定运动强度是有效的,因为利用个体乳酸阈水平为基准进行训练,可抑制肌肉组织中的代谢性酸中毒的早现。Mader等研究者用血乳酸测定了田径、游泳等运动项目的运动强度,用这些值作为基准选择适当的运动强度来指导训练,并使教练员在训练中掌握运动员代谢性酸中毒发展的水平。这种方法既改善了运动员的乳酸阈的水平,同时也提高了运动成绩。由于最大摄氧量受遗传因素影响难以提高,而提高乳酸阈的水平可改善肌肉的有氧能力。因为在这种强度下训练,肌肉的有氧能力的提高不受最大摄氧量的限制。研究表明,乳酸阈时的血乳酸浓度可维持30分钟而不增加,优秀运动员可达到50分钟。因此,在田径、游泳等周期性项目中,利用个体乳酸阈值:安排训练强度已被广泛利用。同时,在足球、手球、橄榄球等项目中利用个体乳酸阈作为评价训练强度的指标,发展运动员有氧耐力的方法也引起了教练员的重视。 3.制定康复健身运动处方 在有些国家,乳酸阈、通气阈已被用于病人的康复和中老年人健身运动处方的制定。 乳酸阈 (LT) 乳酸阈也是反映有氧能力高低的指标.它反映人体在递增负荷运动中,血乳酸浓度未突增前,最大吸氧量实际所利用的百分比,即最大摄氧量的利用率,故有人认为,在反映机体有氧能力上,它比最大摄氧量的价值更大. ①乳酸阈的表示方法与正常范围 通常以乳酸阈时的吸氧量占本人最大吸氧量的百分比来表示.目前对乳酸阈的一些机制和判断标准尚无国际统一规范,因此无法提出乳酸阈的标准正常值.但一般说来,正常成年健康男子的乳酸阈值为55~65%最大摄氧量.据现有文献资料综合来看,总的是有训练者的乳酸阈比无训练者高,耐力运动员的乳酸阈比非耐力运动员为高. ②乳酸阈的推算 乳酸阈的直接测定,需要复杂贵重的实验设备,韦尔特曼经研究提出用3200m跑的时间来推算个人乳酸阈,再用此值除以其O2max,便可得出用 O2max%表示的乳酸阈值. 男子: O2 (ml·kg-l·min-l)=112.0-[5.31×跑3200m的时间(min) ] 女子: O2 (ml·kg-l·min-l)=[-1.123×跑3200m的时间(min)]+47.66 综上所述,最大摄氧量和乳酸阈都是评价有氧能力的指标,可以提供许多关于有氧适能和成绩潜力的信息.有人认为最大摄氧量表明运动强度方面的能力,乳酸阈表明耐力持续时间的能力. |


“肌肉网”

“健身餐食谱”

“教练库”