|
来自:陈守良 《动物生理学》(第二版)北京大学出版社
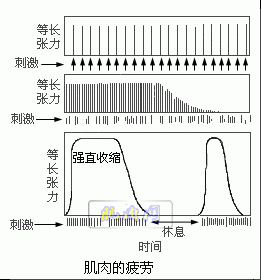
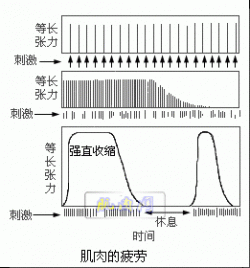
以每秒一次的频率分别刺激蛙的一对缝匠肌,不过对一块肌肉供给氧,而另一块肌肉不供给氧,两块肌肉都随着刺激的频率收缩,在无氧环境中的肌肉较快地停止收缩,在有氧环境中的肌肉维持收缩的时期较长。当两块肌肉都疲劳后停止刺激,休息5分钟,再刺激,供给氧的肌肉恢复收缩能力,而无氧的肌肉不能恢复收缩能力(图示)。
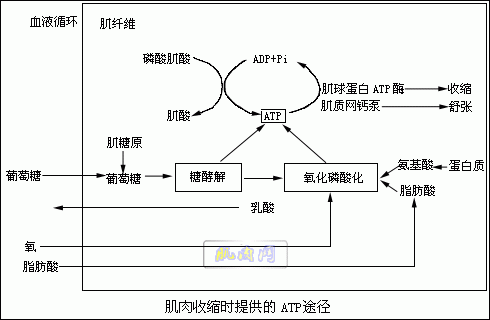
说明肌肉收缩并不一定需氧,而肌肉疲劳后恢复时必需有氧。可见肌肉收缩的直接能源为无氧代谢过程,而恢复所需能源是需氧代谢过程。当肌肉完全疲劳不能收缩时,肌肉中ATP的浓度很低。肌肉恢复收缩能力后,肌肉中ATP的浓度升高。 如果肌肉的收缩只依靠现存的ATP就不可能维持多次收缩,很快就会疲劳。肌肉能维持较长时间的收缩必然能迅速产生ATP以补足由于分解而减少的ATP。有三种途径供给ATP:①磷酸肌酸;②糖酵解;③氧化磷酸化。当肌肉开始收缩时,ATP分解。首先由磷酸肌酸提供合成ATP的能量。CP含有能量和磷酸,可以把一分子的ADP转变为ATP和肌酸(C):
在静息状态,肌肉细胞中ATP的水平较高,促进CP的形成,CP的浓度可达ATP浓度的5倍。肌肉开始收缩,ATP迅速下降时,CP与ADP作用迅速形成ATP,以致ATP的浓度在肌肉开始收缩时下降很少而CP的浓度急剧降低。 如果肌肉收缩活动超过几秒钟,肌肉细胞必须从其他的来源得到ATP的供应。中等水平的肌肉活动时,大部分ATP是由氧化磷酸化的过程提供的。而糖、脂肪和蛋白质则给这个过程提供能源。 当肌肉强烈活动时,ATP很快分解,一系列因素限制了通过氧化磷酸化提供ATP的能力。首先是氧的供应不足,成为限制因素。无氧糖酵解开始提供ATP。虽然需氧的葡萄糖氧化磷酸化过程产生大量的ATP,但速度较低。糖酵解过程产生的ATP数量较少,但速度却高得多。在同一时间内,氧化磷酸化1分子的葡萄糖可产生30分子的ATP,而1分子的葡萄糖通过糖酵解成乳酸可产生2分子的ATP。因此,在激烈的运动时,糖酵解成了提供迅速分解的ATP的另一来源。糖酵解的产物(乳酸)扩散出肌肉组织进入血液中。 肌纤维中的葡萄糖由肌糖原转化而成,激烈的运动把细胞中贮存的肌糖原消耗后就会引起疲劳。在十分激烈的活动中,ATP分解的速度超过了糖酵解可能提供ATP的速度,即使细胞还含有肌糖原也会产生疲劳。图示概括了上述的过程。
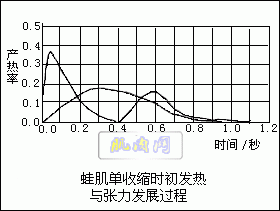
经过一段激烈运动之后,肌肉细胞中产生了一系列的变化:磷酸肌酸的水平降低,大部分肌糖原转变成乳酸。要恢复到原来的状态必须增加糖原的贮备,重新合成磷酸肌酸,这些都需要能量。因此,在肌肉收缩活动停止以后,仍然要消耗氧以供应这些合成过程所需要的能量。运动越激烈,运动时间越长,使肌肉恢复原状所要的时间也越长。肌肉把化学能转变为功的效率约为40%。如果把恢复肌糖原和磷酸肌酸到原水平所需的能量也计入,则总效率只有20%左右,80%的能量转变为热而散发了。 肌肉收缩时的产热问题,早在1848年Helmholtz就已经开始研究,他发现蛙肌强直收缩时温度随即升高。但是深入的研究是在本世纪的上半叶进行的,特别是A.V.Hill的研究确定了伴随肌肉的强直收缩和单收缩所发生的产热过程。Hill用许多热电偶组成热电堆并配以灵敏的电流计可以测量出微小的温度变化,甚至可以记录10-6℃的温度差。 肌肉单收缩时产热过程可以分为两个主要时期,首先在收缩期和舒张期产生初发热,其次在收缩结束后的恢复期产生迟发热。蛙肌作等长单收缩时产生的初发热约每克肌肉0.003卡。初发热的大部分是在肌肉的收缩期产生的,接着产热量减少,在舒张期又增加(图示)。
不论在有氧或无氧条件下,初发热量都相同,这也说明肌肉的收缩与舒张并不一定需氧。在无氧条件下,恢复期的产热量很小,而在有氧条件下,恢复期产热持续几分钟,大体与初发热量相等,叫做需氧迟发热。 |


“肌肉网”

“健身餐食谱”

“教练库”